
 Our visitation days for prospective Physics graduate students at the University of Oregon — people who have been admitted and are deciding whether or not to come — will occur this Friday and Saturday. There’s a poster session, and it occurred to me that we don’t have any posters from my lab that provide a good overview of everything we do. (There’s one lying around from 2013, but it’s severely outdated.) I decided, therefore, to make one myself, as usual severely underestimating how long it would take. It was a good exercise, though.
Our visitation days for prospective Physics graduate students at the University of Oregon — people who have been admitted and are deciding whether or not to come — will occur this Friday and Saturday. There’s a poster session, and it occurred to me that we don’t have any posters from my lab that provide a good overview of everything we do. (There’s one lying around from 2013, but it’s severely outdated.) I decided, therefore, to make one myself, as usual severely underestimating how long it would take. It was a good exercise, though.
Having finished — and having shipped it off in time for printing on cloth — I thought I’d repackage the contents for a blog post, to which I could perhaps point interested students. Of course, I have a web page for the lab as well, but that serves a variety of different purposes and audiences. Before cutting and pasting here, here’s a low-res image of the entire poster (including an embarrassing typo in the first line…) :
Microbes, Membranes, Microscopes, and More
An overview of the Parthasarathy Lab
Department of Physics, The University of Oregon

Introduction: Biophysics
Our lab explores biophysics – asking and answering questions about how physical laws are manifested in the living world, how the physical properties of biological materials guide and constrain life, and how living systems are organized in time and space. We primarily investigate lipid membranes (the major component of cellular membranes) and gut microbial communities (vast, dynamic, and mysterious ecosystems contained inside each of us). We develop and implement new experimental tools, such as 3D microscopes, and computational tools, such as cutting-edge machine learning methods, to make these studies possible. Most of our experiments are unique, pushing the boundaries of what has been seen or studied, which we find exhilarating!
Membranes
Cellular membranes are remarkable materials: flexible, two-dimensional fluids. Their flexibility is important to the control of shape, as membranes are pinched off into cargo packets, as organelles are formed within cells, and as cells themselves divide. Their fluidity is important to the ability of membrane proteins – encoded by over a third of the genes in the human genome – to move and associate with one another as they build structures and perform chemical reactions.
The lipid bilayer forms the underlying architecture of all cellular membranes, and determines the membrane’s physical properties. We perform experiments to quantify characteristics such as fluidity and rigidity, often developing new methods involving microscopy, micromanipulation, and particle tracking.
A giant vesicle – a spherical shell of lipid bilayer.
This membrane [movie, above] has “phase separated” into cholesterol-poor (orange) and cholesterol-rich (dark) regions, both of which are two-dimensional liquids. From the motion of the dark domains, we can infer physical properties of the membrane, such as its viscosity.


Applying tension to vesicles. Schematic (above, top) and actual image (above, bottom) of a giant vesicle, with mechanical tension applied by a micropipette. Scale bar: 20 μm. With this approach, we were able to quantify the relationship between tension and fluidity in membranes. The three-dimensional analogue of this (the pressure-viscosity relationship in liquids) has been studied for over a century, but ours was the first measurement of this in (2D) membranes!
Suggested Parthasarathy Lab papers about membranes:
- V. L. Thoms, T. T. Hormel, M. A. Reyer, R. Parthasarathy, “Tension Independence of Lipid Diffusion and Membrane Viscosity.” Langmuir. 33, 12510–12515 (2017). [Link]
- Tristan T. Hormel, Matthew A. Reyer, Raghuveer Parthasarathy, “Two-Point Microrheology of Phase-Separated Domains in Lipid Bilayers.” Biophys. J. 109: 732-726 (2015). [Link]
- Tristan T. Hormel, Sarah Q. Kurihara, M. Kathleen Brennan, Matthew C. Wozniak, and Raghuveer Parthasarathy, “Measuring Lipid Membrane Viscosity Using Rotational and Translational Probe Diffusion,” Phys. Rev. Lett. 112, 188101 (2014). [Link]
- Andrew F. Loftus, Sigrid Noreng, Vivian L. Hsieh, and Raghuveer Parthasarathy, “Robust Measurement of Membrane Bending Moduli Using Light Sheet Fluorescence Imaging of Vesicle Fluctuations,” Langmuir 29: 14588–14594 (2013). [Link]
Microbial Communities
You, like all animals, are not a single species. Each of us is host to tens of trillions of microbes, mostly resident in the intestines, whose profound importance to health, disease, and development is just beginning to be understood. Conventional methods, based mainly on DNA sequencing, tell us what species or genes are present, but provide little or no insight into their physical structure or temporal dynamics – things that must be important for the functioning of a multispecies ecosystem. How do bacterial colonies nucleate and grow? How (and why) do bacteria swim in a complex environment? How do different groups compete? What does “equilibrium” look like in a complex system of active, strongly interacting agents?

A larval zebrafish. We study the gut microbiome inside this transparent, tractable model vertebrate. (Bar: 250 μm; red dye indicates the intestine)
Ours are the first images and movies of gut microbes inside a vertebrate gut!
We find different spatial structures for different species. This Vibrio bacterium lives as highly motile individuals (Movie below; bar: 50μm).
Watching multiple species reveals mechanisms of competition. This Vibrio population grows continuously, but another native bacterium, Aeromonas, shows sudden, orders-of-magnitude collapses well described by stochastic dynamical models. We’ve shown that these collapses are driven by the physical activity of the gut and the structure of the different bacterial communities. (Movie below, spanning several hours. Bar: 200 μm).
How do disease-causing bacteria invade the gut microbiome? Can we “reboot” gut microbial communities by harnessing certain species’ ability to kill other cells?
To explore these questions, we examine Vibrio cholerae, the pathogen that causes cholera, a disease that infects millions of people and kills tens of thousands every year.
Vibrio cholerae’s “Type VI Secretion System” (T6SS) acts as a syringe with which the bacterium stabs adjacent cells and delivers toxins (sketched above).
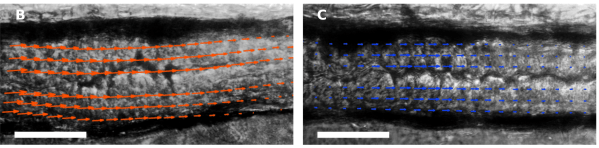
We watched Vibrio cholerae with and without the T6SS attempt to invade the zebrafish gut. We found that the T6SS can control the pathogen’s ability to displace a native microbial species, and moreover that it does so by altering the host’s mechanical contractions of the gut – a previously unknown ability of bacteria to manipulate animal physiology. Shown: image velocimetry of intestines invaded by Vibrio cholerae with (above left) and without (above right) the T6SS.
Suggested Parthasarathy Lab papers about Microbial Communities:
- S. L. Logan, J. Thomas, J. Yan, R. P. Baker, D. S. Shields, J. B. Xavier, B. K. Hammer, R. Parthasarathy, “The Vibrio cholerae Type VI Secretion System Can Modulate Host Intestinal Mechanics to Displace Commensal Gut Bacteria.” Proceedings of the National Academy of Sciences (2018). [Link]
- M. J. Taormina, E. A. Hay, R. Parthasarathy, “Passive and Active Microrheology of the Intestinal Fluid of the Larval Zebrafish.” Biophysical Journal. 113, 957–965 (2017). [Link]
- Travis J. Wiles, Matthew L. Jemielita, Ryan P. Baker, Brandon H. Schlomann, Savannah L. Logan, Julia Ganz, Ellie Melancon, Judith S. Eisen, Karen Guillemin, Raghuveer Parthasarathy , “Host Gut Motility Promotes Competitive Exclusion within a Model Intestinal Microbiota.” PLoS Biol. 14(7): e1002517 (2016). [Link]
Microscopy
Our biophysical investigations are all based on optical microscopy, often pushing the boundaries of what can be imaged. All of our studies of gut microbial communities, and some of our membrane work, use light sheet fluorescence microscopy, a technique in which a thin sheet of fluorescence excitation light optically sections a specimen. This approach enables fast, high resolution, three-dimensional imaging with low phototoxicity.



Our home-built light sheet microscopes are ideal for studies of the microbiome. We’ve recently created an automated microscope capable of high-throughput imaging of live specimens. In general, we believe in adopting cutting edge techniques to perform new types of scientific investigations.
Machine Learning, Image Analysis
Our experiments generate large amounts of data, leading to the challenge of generating quantitative insights from images. For example, we wish to track membrane-anchored particles with nanometer precision, and identify bacteria in complex, 3D images in which the population can span five orders of magnitude. To tackle this, we develop and implement advanced computational approaches, including machine learning methods like neural networks.

Top: Gut bacteria imaged at 1.6 hrs. post-inoculation. Few bacteria are present. Inset: a single bacterium. White bars show autofluorescent zebrafish sources. Bottom: The same fish 7.5 hrs. later has a large bacterial population. Bars: 100 μm.
People
Our lab is made of a wonderful team of graduate students, undergraduates, and postdocs. Group photo, Summer 2017:

More Photos: Philip Jahl conquers Jenga, Savannah Logan conquers Smith Rock, and Vince Thoms makes crème brûlée at lab meeting.



We also have an excellent set of collaborators, including the lab of Karen Guillemin (U. of Oregon, Biology), many other labs at UO, and Brian Hammer (Georgia Tech) and Joao Xavier (Memorial Sloan-Kettering).
We’ve been supported by the NSF, the NIH (see meta.uoregon.edu), Research Corporation for Science Advancement, the Gordon and Betty Moore Foundation, the Simons Foundation, and the Kavli Foundation.
Email: raghu@uoregon.edu Group web page: http://pages.uoregon.edu/raghu/
Today’s illustration: Part of a drawing of a potassium channel protein. I might post the rest of it, and describe it more, in a future post.
— Raghuveer Parthasarathy, April 2018
