

About a paper from my lab [1] on competition and cooperation among gut microbes.
Is the whole more than the sum of its parts? This question arises throughout the sciences, as one wonders whether understanding the constituents of some system suffices to understand the system as a whole. Sometimes the answer is “yes.” Electromagnetic fields, for example, are additive in simple media — each new piece adds to the others in a straightforward way. Sometimes the answer is “no.” Studying a single iron atom won’t reveal that spontaneously magnetic materials exist, and two grains of sand don’t explain a sandpile. As Philip Anderson famously said, “more is different.”
This question comes up in many ecological contexts: one wonders whether interactions between species in an ecosystem can be understood as a combination of interactions between pairs of species. Does knowing, for example, how A & B, B & C, and A & C interact allow us to predict what happens when A, B, and C are all together? If it doesn’t, there must be “higher order” or non-pairwise interactions. The existence of higher order interactions is difficult to determine, both in terms of gathering the requisite experimental data as well as interpreting it. All this applies to microbial ecosystems as well as those containing larger creatures, and studies assessing higher order interactions among species of microbes date back to at least the 1960s and continue to the present [2, 3]. There have been a few such studies involving animal-associated gut microbiomes — excellent investigations of the bacterial residents of fruit flies [4] and nematode worms [5] — but none in any vertebrate, where the complexities of anatomy and specific selection of symbiotic microbes are greater.
The topic of interaction rules for gut bacteria is of more than abstract interest. Gut microbes influence many aspects of health and disease, motivating a lot of interest in the question of how one might reshape intestinal communities. To do this rationally, one would like to know how different species interact, for example if one wants to introduce a new, “helpful” species that one hopes will flourish in the gut. Given N species, is understanding the N(N-1)/2 pairwise interactions sufficient to characterize every possible subset, or is every subset special?
We decided, therefore, to explore interactions between bacterial species in the zebrafish gut. As I’ve written about before, zebrafish are an amazing model organism that make possible explorations of all sorts of processes at play in vertebrates, including humans, and my lab has been using larval zebrafish to illuminate the biophysics of the gut microbiome for several years. (See, for example, here, and here.) Zebrafish can be derived devoid of any gut microbes, after which one can expose them to specific sets of species, allowing controlled experiments on inter-species interactions. We’ve explored this before just with pairs of species (for example, this and this); could we increase the complexity?
Okay, cats, we rumble! [6]
We worked with five different species of bacterial native to the zebrafish gut. “We” is, of course, mainly students in my research group, most notably Dylan Martins, Teddy (Edouard) Hay, and Deepika Sundarraman. Dylan was a very hard-working undergraduate who graduated last year. Teddy was a graduate student, who has since graduated and started his own company making medical diagnostics. Deepika is currently a graduate student in the group, now pushing this work in new directions involving imaging the interactions among these and related species and also between bacteria and the host immune system — I’m sure I’ll describe her amazing images and insights in a future post! For now, I’ll focus on our 2020 paper, which has the unusual distinction of being our only paper on the gut microbiota that does not involve any microscopy.
We first pitted pairs of bacterial species against each other, all ten pairwise combination of the five species, assessing the abundance of each in the gut if allowed to simultaneously colonize bacteria-free zebrafish. We examined hundreds of larval fish, delicately removing their intestines by dissection under a microscope. (Dylan excelled at this. I’ve done it only a few times myself, not well.)
Some pairs barely affected each other, with each species as abundant with the other present as it would be on its own, on average. (Blue, in the illustration below for example, is barely affected by the presence of Pink.)
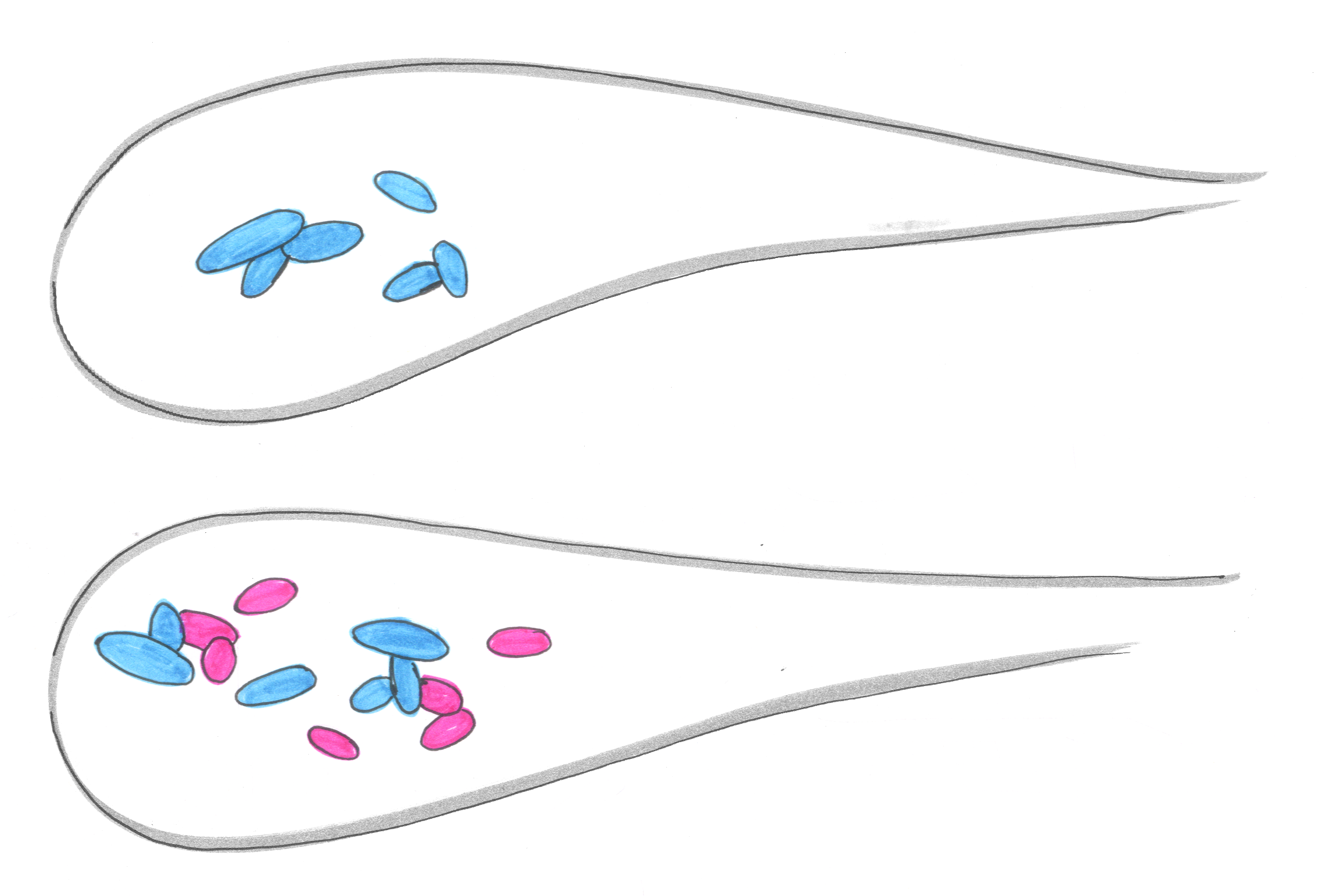
There was one mildly cooperative interaction — a species whose population was slightly enhanced by the presence of another. Most notably, there were several strongly negative interactions, in which a species suffered if a particular rival were present. (Green in the illustration below, for example, is pummeled by pink.) Our approach didn’t reveal what the the interactions are, and some of our current work aims to observe the fights between the rival gangs for the turf of the gut.
Tidepools, revisited.
Next, we tackled the question of how gut bacterial species interact if other species are present.
Inferring the influence of any particular species on others in an ecosystem is in general challenging. In a classic, clever experiment that took place here in the Pacific Northwest in the 1960s, ecologist Robert Paine visited tide pools, home to an array of wonderful creatures like sea urchins, anemones, barnacles, and muscles, and systematically tossed out the starfish, peeling them from the rocks and flinging them as far as he could. He removed every starfish. (It may seem a bit heartless, but it was only a 25 foot span of the shore, and it taught us a lot about ecology.) Then, over the following months and years, he surveyed the populations of the remaining tide pool inhabitants, uncovering a steep drop in the pool’s diversity, as species like mussels, unchecked by starfish predation, took over large swathes of rock. Based on this and other studies, Paine conceived of the notion of “keystone species” that have a large impact on their entire ecosystem. For a nice article by Sean B. Carroll on Paine’s work, see here; it’s an excerpt from his The Serengeti Rules: The Quest to Discover How Life Works And Why It Matters a popular science book by the same publisher as my upcoming book! For a nice video reenactment of Paine’s starfish hurling, see here.
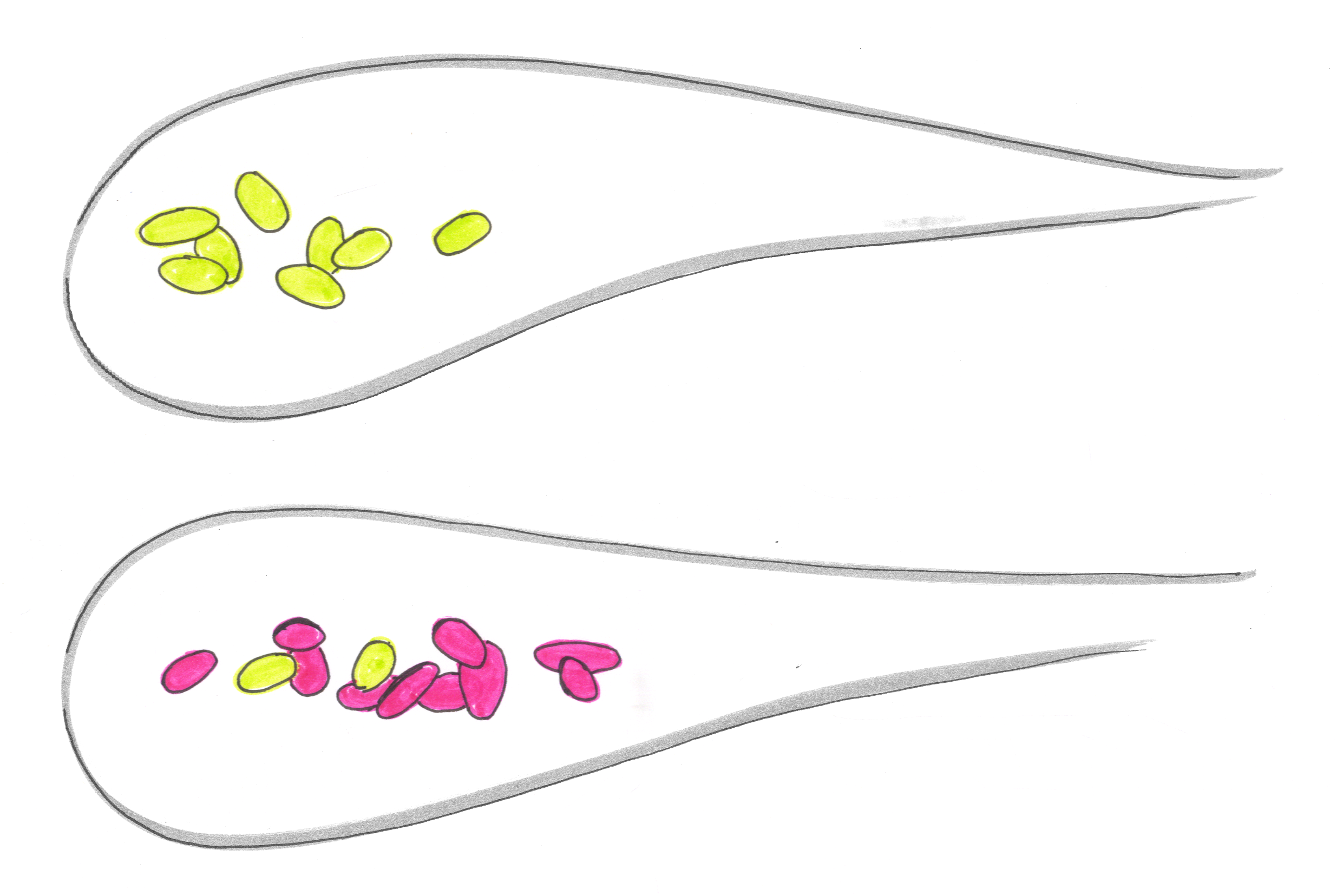
What we would have liked to do is the zebrafish gut analog of Paine’s experiment, removing one species and seeing what happens to the others. (Years ago, I already likened your gastrointestinal tract to a tidepool; I’m now doing it again for totally different reasons!) There’s no good way to perform the removal, however. Instead, we took a different route, comparing fish colonized by four bacterial species to fish colonized by five. The difference in abundance of a species in the two cases reveals its interaction with the fifth species amid the backdrop of the others. In the illustration, for example, pink no longer suppresses green, when orange, blue, and purple are present.
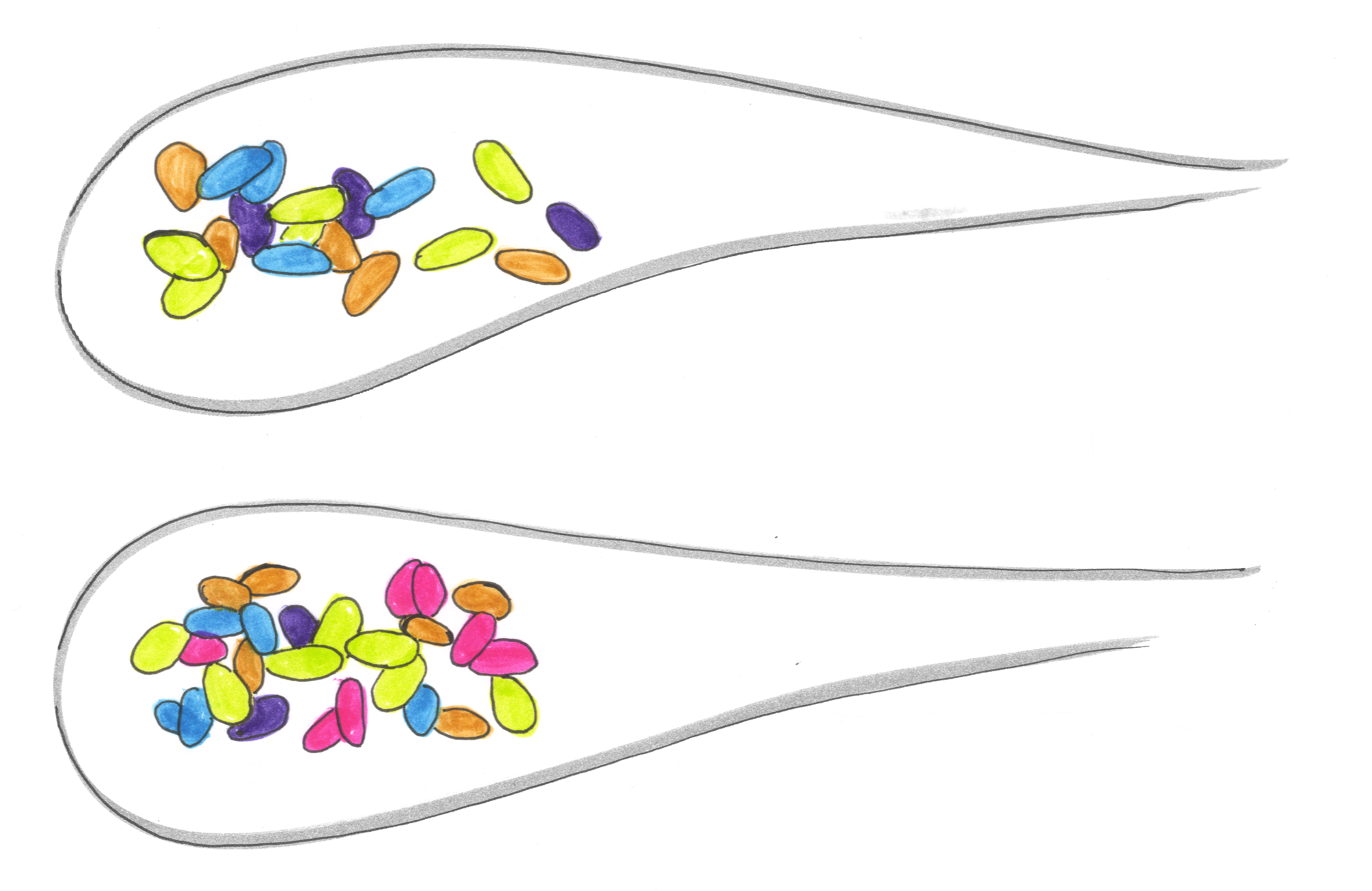
Again averaging over many fish, what we found was that the strong negative interactions of our pairwise competitions were greatly diminished in the multi-species case. The case sketched in the cartoon above held quite generally. Similarly, if we used the pairwise data to try to predict the abundance of each species in five-species communities, the predictions failed dramatically — communities were home to more, and more equally distributed, species than we would have expected.
Our results supply the first evidence that higher order interactions are substantial in a vertebrate gut, and that they act to dampen strong competition and stabilize diverse multispecies communities. Intriguingly, in contrast to conventional wisdom dating to ecological theory of the 1970s, a lot of fascinating recent theoretical work, much of it done by physicists, suggests that higher-order interactions can quite generally promote coexistence [7-9]. In this context, our findings are perhaps not as shocking as they might at first seem. The failure of pairwise interactions — relatively easy to measure — to describe multi-species communities is perhaps discouraging from the perspective of microbiome engineering. On the other hand, the robust coexistence of bacterial species in the gut suggests general principles at play, likely relevant to your intestines as well as the zebrafish’s.
Our assessment of inter-species interactions took a huge amount of work, and only begins to answer the question of how gut bacterial compete and cooperate. Why does having multiple species foster stability? Do bacterial colonies, by virtue of their three-dimensional structure, shield each other from competitive interactions? (We’re of course fond of this physics-centered interpretation!) Do different species form networks of nutrient exchange? The answer to these questions will, I hope, point to ways of understanding, and manipulating, the gut microbiome.
Today’s illustration.
A starfish on a rock. (What else could I do?)
— Raghuveer Parthasarathy; March 18, 2021
Notes
[1] Deepika Sundarraman*, Edouard A. Hay*, Dylan M. Martins*, Drew S. Shields, Noah L. Pettinari, and Raghuveer Parthasarathy, “Higher-Order Interactions Dampen Pairwise Competition in the Zebrafish Gut Microbiome.” mBio 11: e01667-20 (2020). Link
[2] J. H. Vandermeer, “The Competitive structure of communities: an experimental approach with protozoa.” Ecology 50: 362-371 (1969). Link
[3] H. Mickalide, S. Kuehn, “Higher-order interaction between species inhibits bacterial invasion of a phototroph-predator microbial community.” Cell Syst. 9: 521-533 (2019). Link
[4] A. L. Gould, V. Zhang, L. Lamberti, E. W. Jones, B. Obadia, N. Korasidis, A. Gavryushkin, J. M. Carlson, N. Beerenwinkel, W. B. Ludington, Microbiome interactions shape host fitness. PNAS 115: E11951-E11960 (2018). Link
[5] A. Ortiz, N. M. Vega, C. Ratzke, J. Gore, Interspecies bacterial competition regulates community assembly in the C. elegans intestine. The ISME Journal, 1-15 (2021). Link
[6] West Side Story, https://www.moviequotedb.com/movies/west-side-story/quote_29495.html
[7] A. Posfai, T. Taillefumier, N. S. Wingreen, Metabolic Trade-Offs Promote Diversity in a Model Ecosystem. Phys. Rev. Lett. 118, 028103 (2017).
[8] J. E. Goldford, N. Lu, D. Bajic, S. Estrela, M. Tikhonov, A. Sanchez-Gorostiaga, D. Segrè, P. Mehta, A. Sanchez, Emergent simplicity in microbial community assembly. Science. 361, 469-474 (2018).
[9] R. D’Andrea, T. Gibbs, J. P. O’Dwyer, Emergent neutrality in consumer-resource dynamics. bioRxiv, 710541 (2019).
